Page created on April 4, 2018. Last updated on December 18, 2024 at 16:55
B-cell subsets and their characteristics
The B1 type B-cells are, like the γδ T-cells, made only during the embryonic life. Instead of dying out after that however, they renew themselves in our body, outside of lymphoid organs, in the periphery. The presence of a spleen is essential in the survival of the B1 B-cells. They produce low-affinity antibodies and sometimes even antibodies that can bind to self-antigens (antigens of proteins that are normally found in the body), so-called autoantibodies (auto means self in Greek). They can be found in the serosa of body cavities. They are highly positive for IgM.
The B2 type B-cells are divided into two types, the marginal zone B-cells and the follicular B-cells. They originate from the bone marrow and mature in the spleen.
The marginal zone B-cells can be found in the marginal zone of the spleen (and not the lymph node), where they border the red pulp. They are also highly positive for IgM. They don’t migrate anywhere else but stay where they are, meaning they are sessile. They also express many complement receptors. They constitute most of the T-cell independent antibody response.
The follicular B-cells are the ones that are found in blood and make up the follicles in the spleen and in the cortex of lymph nodes. They are highly positive for IgD and can easily migrate and recirculate. They are also what constitutes most of the T-cell dependent antibody responses, as we will see below.
A special type of T-helper cells called follicular T-helper cells or TFH are important in this topic. They differentiate from T-helper cells when they have protein antigen presented to them, and express high levels of the chemokine receptor CXCR5, which will draw them into the follicles.
What makes B-cells start producing antibodies?
B-cells must stay alive in the periphery until they meet with an antigen they can bind to. A protein called BAFF is important in this survival of the B-cells.
B1 B-cells constantly produce antibodies. When activated, they move from the serosa to the lamina propria of the gut. Not much else is known about them.
B2 B-cells can have two responses to an intruder, and the response depends on the antigens of the intruder. If the antigen is a protein, the binding between B-cell and the protein antigen is not a strong enough signal for the B-cell to begin an immune reaction. In this case, a T-cell is essential to give a stronger signal to the B-cell, to start the immune reaction. That is why the T-cell dependent immune response only happens in response to protein antigens.
The interaction between B-cell and follicular T-helper cell begins only after the B-cell has bound the antigen by itself, and the follicular T-helper cell has had the antigen presented to it by an APC. The B-cell will phagocyte the antigen and present antigen peptides on MHC II. The TCR on the TFH-cell will bind to the MHC II on the B-cell, and the CD40 on the B-cell will bind to CD40L on the T-cell. The T-cell will also produce cytokines, that will bind to cytokine receptors on the B-cell. This will activate both cells. More on this further down.
In the case of antigens that are not proteins, like lipids or carbohydrates, the immune response is T-cell independent, which means that T-cells are not needed for the reaction. This is because a pathogen often has many copies of the same epitope on the surface in the case of non-protein antigens, and many of these epitopes can bind to separate BCRs on one B-cell. When many BCRs are stimulated at the same time, the signal is strong enough for the B-cell to initiate the immune response without the help of a T-cell.
The extrafollicular reaction (or plasmacellular reaction) begins almost immediately after the initial infection. It happens outside the follicles (hence the name), and is T-cell independent, but also happens during the T-cell dependent response. It’s the body’s initial reaction to an intruder and causes marginal zone B-cells to differentiate into plasma cells to produce low-affinity IgM. In the case of protein antigens, it’s a sort-of temporary defence while the body waits for the germinal centre reaction to start producing high affinity antibodies. In the case of non-protein antigens, it’s the only immune response the body will have to the antigen. The extrafollicular reaction does not cause formation of memory B cells.
The germinal centre reaction is protein-antigen dependent. It begins 7 days after the initial infection, so it is delayed compared to the extrafollicular reaction, and is done mostly by follicular B-cells. It will generate a germinal centre in the secondary lymphoid organs. It happens inside the follicles and is dependent on many cellular components and is T-cell dependant. The goal of the germinal centre reaction is the production of plasma cells that secrete antibodies with high affinity towards the antigen, and to produce memory cells that can make the response to the same antigen in the future quicker. The germinal centre reaction mostly ends up producing IgM and IgG antibodies, but can also produce antibodies with other isotypes, depending on the antigen.
Follicular dendritic cells (FDCs) are also important. These cells are not related to dendritic cells (despite their name), and are not even hematopoietic in origin, but come from mesenchyme. They don’t have phagocytic activity and are non-adherent. They depend on an extracellular protein called LTβ to differentiate, and also on TNF to migrate to the correct tissue and location. They express VCAM-1, an adhesion molecule used to bind centrocytes (they’ll be defined later). Lastly, FDCs can store antigens for a long time, as part of an immune complex called the iccosome. The point of this will be clear in the next topic.
Both types of B2 B-cells are involved in the germinal centre reaction. The marginal zone B-cells will produce IgM and transport the antigen to the follicular B-cells with the help of the complement system, a system we will talk about more in a later topic.
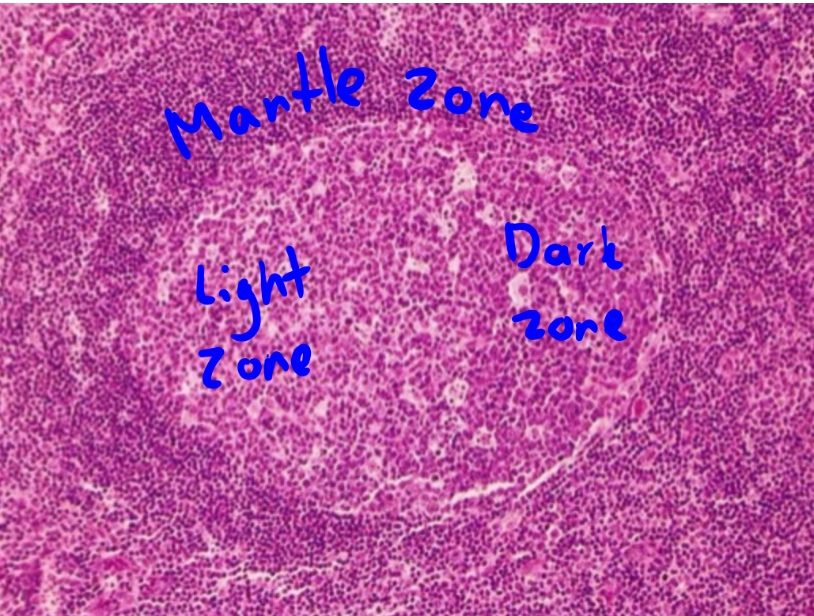
The binding of the TFH and B-cell during the T-cell dependent response activates both cells, and causes the B-cell to start dividing, creating clones of itself with the same antibody-producing properties. Some of these clones will remain outside the follicle and become plasma cells that will produce low affinity antibodies. This is the extrafollicular reaction. Some of the clones will migrate into the lymphoid follicle, where they will start to rapidly divide. This takes place in the germinal centre. Some of these clones will undergo isotype switching, which we will learn more about in the next topic. The rapidly dividing B-cells are now known as centroblasts and make up the dark zone of the germinal centre. The centroblasts express a transcription factor called Bcl-6, and undergo affinity maturation, which we will also talk about in the next topic. Affinity maturation increases the affinity of the antibodies to the antigen. A cell type called tingible body macrophages will phagocyte cells that are not formed correctly during the division. They are also found in the dark zone.
After some rounds of cellular division, some centroblasts will become centrocytes and create the light zone of the germinal centre.