Page created on April 22, 2018. Last updated on November 19, 2018 at 17:16
Suppression of the immune system at the cellular level
There are many mechanisms involved in suppressing the immune response to make sure it doesn’t go haywire. We’ll start with the suppression mechanisms on the cellular level.
After antigen-presenting cells have phagocytosed a pathogen or cell, they will be inhibited. This is due to the content of thymidine (T) in any organism containing DNA. The ingested thymidine inhibits the proliferation of the APCs, which makes sure that the APCs only migrates to lymph nodes and don’t start proliferating unnecessarily.
The Treg cells are obviously important in suppression, considering their name. Their phenotype is CD4+/CD25+/FoxP3. They have multiple mechanisms for suppressing the immune response. Treg cells have CD25, also known as the IL-2 receptor. Because IL-2 is a cytokine that is essential for the proliferation and differentiation of T-cells. The Treg cells deprive other T-cells of IL-2 by binding to IL-2 themselves, which reduces the amount of IL-2 present, which kills T-cells due to cytokine deprivation.
Both activated T-cells and Treg cells express CTLA-4, a surface adhesion molecule that binds to CD80 and CD86, which are present on APCs. This binding inhibits the APCs and the activated T-cells, reducing their activity. Treg cells also induce the extrinsic apoptotic pathway in effector T-cells with the help of granzymes and perforin. Lastly, Treg cells produce inhibitory cytokines like TGFβ, IL-35 and IL-10, which inhibit the activity of effector T-cells.
We know that the presence of viruses or intracellular bacteria causes dendritic cells to start producing IL-12, which differentiate Th cells to Th1 cells, and parasites cause them to produce IL-4, which causes Th to differentiate to Th2 cells. However, the presence of no pathogen causes dendritic cells to produce IL-10 and TGFβ, which induce the differentiation of T-cells to Treg cells, which inhibits the immune response further.
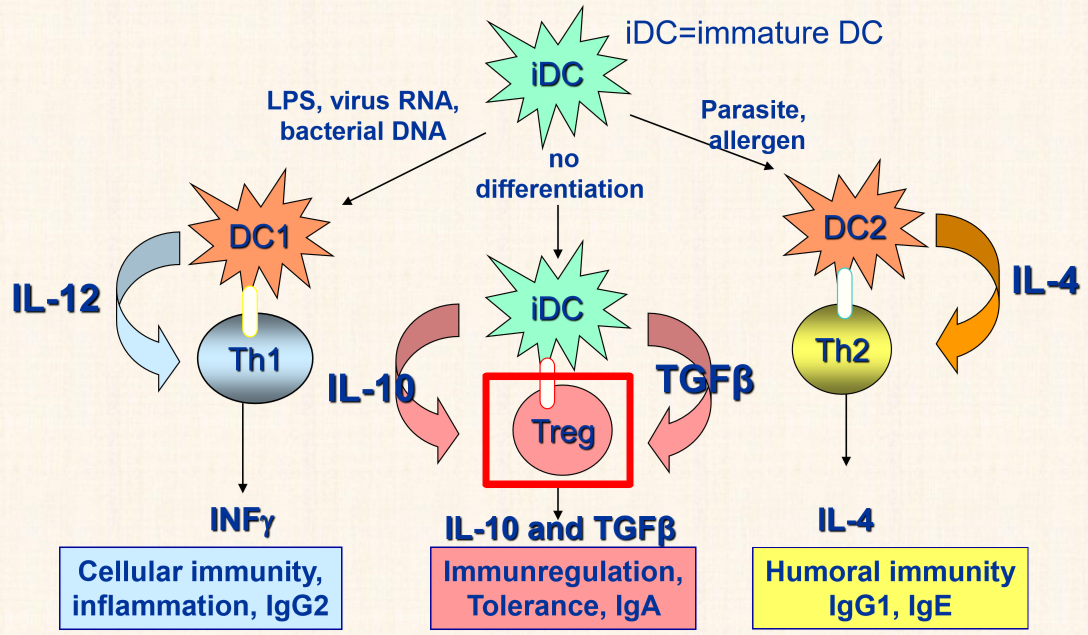
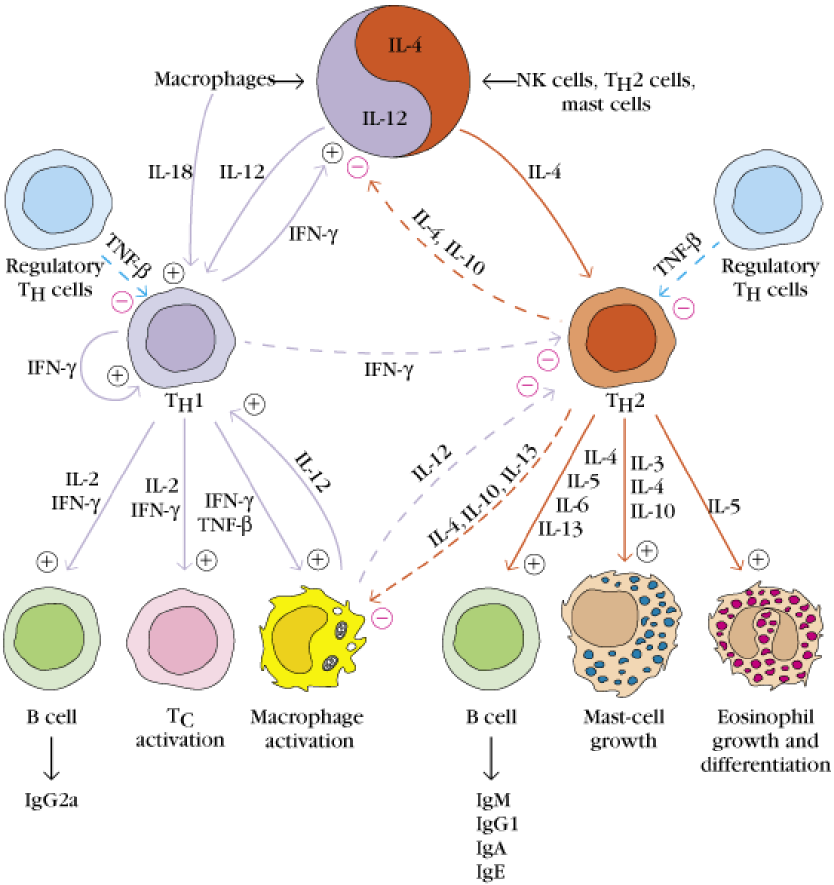
The last suppression of T-cells is the reciprocal inhibition of T helper cells subsets Th1 and Th2. Th1 cells produce many cytokines, among them is IFN-γ, which activates other Th1 cells, but also inhibits Th2 cells. This causes a balance between the two subtypes.
B-cells have a surface protein called CD32, better known as the FcγR II. Recall from topic 13 that this is the only FcγR that has an inhibitory effect on the B-cell. The FcγR II has low affinity for IgG antibodies, so only a large number of antibodies can activate them, and they can only be activated by IgGs that have already bound an antigen. This causes the B-cells to be inhibited in response to an excess of antibodies, so that the B-cells “know” when enough antibodies have been produced.
Suppression of the immune system at the humoral level
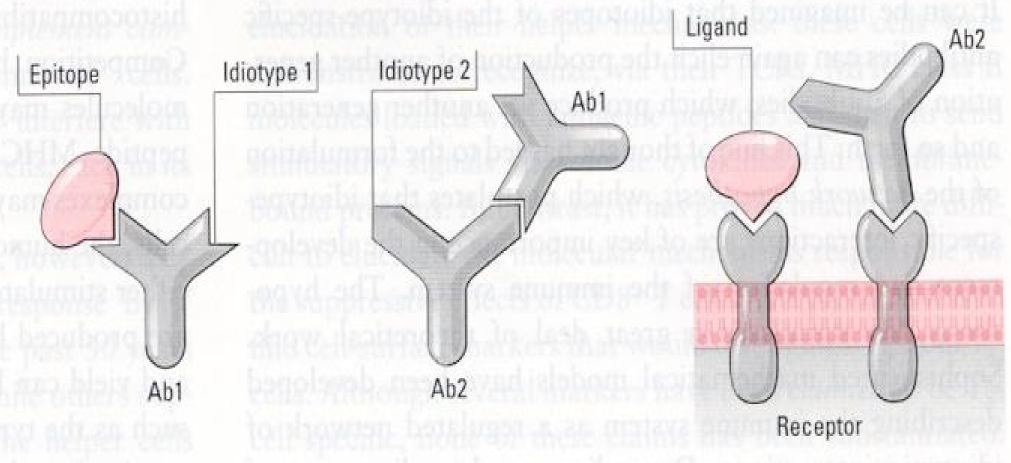
We all know that antibodies bind epitopes of antigens. This binding happens through electrical forces, the binding of positive charges on the antibody to negative charges on the epitope and vice versa. If we think of the epitope as a particle, the antigen-binding site of the antibody that binds that epitope must be like the “mirror image” of the epitope.
Something to note is that the antigen-binding site of every antibody is a foreign structure to the body, because the body has never seen it before, because it is formed from random recombination of genes. This causes the body to produce antibodies against another antibodies’ antigen-binding sites! These antibodies are called anti-idiotype antibodies, because they recognize another idiotype of antibodies. The body can produce anti-idiotype antibodies that bind to a primary antibody to reduce the number of them in the body.
The consequence of this and the “mirror image”-theory is that the body produces antibodies, the antigen-binding sites of which are similar in structure to the original epitope! The figure below explains this better as it can be difficult to understand.
While the body can produce anti-idiotype antibodies, that bind antigen-binding sites of other antibodies, it can also produce antibodies against the other parts of antigens. These are called anti-isotype antibodies and bind to a specific isotype of antibodies, or more specifically the constant regions of the antibodies. Anti-isotype antibodies can be anti-IgG for example. A disease marker called Rheumatoid factor is actually an IgM type antibody that binds to the constant regions of IgG antibodies (anti-IgG antibodies).
An important conclusion to draw from this topic is that immune suppression is not a feature of just one cell type or one mechanism, but by a network of many cell types and regulatory mechanisms.