Table of Contents
Page created on November 11, 2019. Last updated on January 24, 2022 at 16:17
The glomerular filtration rate is approx. 100 mL/min, which equals approx. 180 litres per day. With 180 litres of plasma being filtered into the Bowman space every day you’d expect us to urinate 180L every day. However, we only produce approx. 1,5L of urine every day, and not 180L. The difference of 178,5 litres is reabsorbed by the tubules after filtration.
After the ultrafiltrate leaves the Bowman space it travels through the tubular system before it reaches the collecting duct. While traveling through the tubular system 99% of the ultrafiltrate is reabsorbed back into the peritubular capillaries and returned to the systemic circulation.
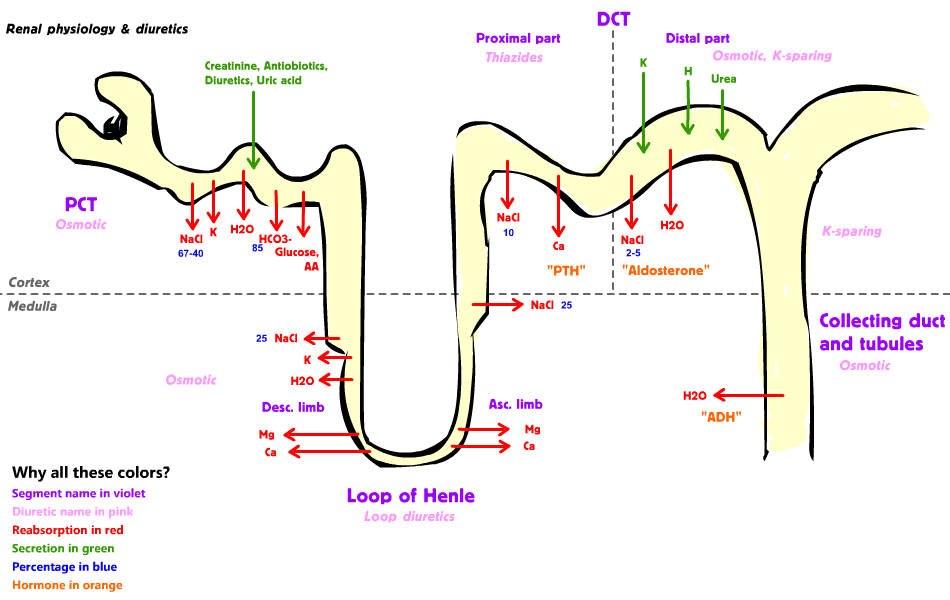
This figure shows the different parts of the tubular system and which substances are secreted and reabsorbed and where. From Wikipedia.
Mechanisms of reabsorption and secretion
For a substance to be reabsorbed it must be transported across the tubular epithelium and into the renal interstitium, and then into the peritubular capillaries. Before we get into the specifics, we need to look at the different mechanisms of reabsorption and secretion the tubular system utilizes. The most important mechanisms are:
- Passive transport
- Active transport
- Primary active transport
- Cotransport/secondary active transport
- Endocytosis
- Pinocytosis
Passive transport:
Passive transport does not require energy. Substances simply move down their concentration gradient (from areas of high concentration to areas of low concentration) or down their electrochemical gradient (towards areas with the opposite charge). Substances pass through cells through specific transport proteins, which are often specific for a certain substance.
Water is always transported passively, by a mechanism called osmosis. Osmosis causes water to diffuse from areas with lower osmolarity to areas with higher osmolarity. A good rule of thumb is that water will always follow Na+. If Na+ is transported in one direction, water will follow.
Active transport:
Active transport requires energy in the form of ATP. Active transport is needed when substances need to move against their concentration or electrochemical gradient. We can distinguish two types of active transport.
Primary active transport occurs when the active transport is directly coupled to ATP. An example is the sodium-potassium ATPase transporter. This transported hydrolyses one ATP into ADP and exchanges one sodium for one potassium.
Secondary active transport or co-transport occurs when the active transport isn’t directly coupled to ATP. This is more complicated as it involves two substances and two transport proteins. The best example is the reabsorption of glucose, which is detailed below.
Endocytosis and pinocytosis:
Pinocytosis is a form of endocytosis, where cells “swallow” large molecules instead of transporting them through transport proteins. Pinocytosis is used to reabsorb proteins.
Symporters and antiporters:
A symporter is a transport protein which transports two or three substances in the same direction simultaneously. An antiporter on the other hand transports multiple substances in the opposite direction.
Proximal convoluted tubule
The proximal convoluted tubule (PCT) is the first part of the tubular system. Most of the ultrafiltrate is reabsorbed here. The most important functions of the proximal convoluted tubule are:
- Reabsorption
- All glucose
- All amino acids
- Uric acid
- 2/3 of all filtered Na+
- 2/3 of all filtered K+
- Secretion
- NH3
- H+
- HCO3– (most of it)
- Creatinine
Reabsorption of sodium:
The majority of sodium which is reabsorbed in the PCT is reabsorbed by active transport. This active transport is mediated by a transport protein called Na+/H+ ATPase, an antiporter which pumps Na+ into the cell and H+ into the filtrate.
Reabsorption of glucose and amino acids:
Reabsorption of glucose and amino acids is mediated by secondary active transport. There is more glucose inside the tubular epithelial cells than in the filtrate, so active transport is required to reabsorb it. This is achieved with the use of a transport protein called sodium glucose co-transporters (SGLT). These proteins are symporters, so they transport sodium and glucose simultaneously. Glucose has to move against its concentration gradient, but sodium has to move with its electrochemical gradient. The force of the electrochemical gradient of sodium is larger than the force of the concentration gradient of glucose, so the net effect is that both sodium and glucose are transported into the cell simultaneously.
The same principles apply to amino acids as well, but they use different proteins.
Thin descending limb of the loop of Henle
The main function of the loop of Henle is to create an osmotic gradient in the kidney. This means that the osmolarity of the kidney interstitium is higher in the deeper parts of the medulla. This is essential for the kidney to concentrate and dilute urine, as we will see in topic 77.
The thin descending loop of Henle is impermeable to Na+, meaning that no Na+ is reabsorbed or secreted here. It is permeable to water, though. The interstitium around the thin descending loop is hyperosmotic, so water will be reabsorbed. Because the filtrate loses water but not Na+ the osmolarity of it increases. The filtrate reaches its highest concentration at the bend of the loop.
Thin ascending limb of the loop of Henle
The thin ascending limb of the loop of Henle is impermeable to water but permeable to Na+. Na+ reabsorption occurs passively. The filtrate becomes less hyperosmolar.
Thick ascending limb of the loop of Henle
The thick ascending limb of the loop of Henle is also impermeable to water and permeable to Na+. Unlike the thin ascending limb however, Na+ reabsorption is active here. Na+, K+ and Cl– are actively reabsorbed by the Na-K-Cl cotransporter, which is a symporter.
As Na+ is continuously pumped out of the filtrate and into the interstitium the filtrate becomes less and less osmotic. The osmolarity of the interstitium also decreases toward the cortex, because of this. At the end of the thick ascending limb the filtrate is iso-osmolar.
Distal convoluted tubule
The macula densa lies in the first part of the distal convoluted tubule (DCT). The DCT can be divided into the proximal part and the distal part.
Proximal part:
The permeability of the proximal part of the DCT is similar to that of the thick ascending limb, in that they’re both impermeable to water and that ions are reabsorbed in both. Calcium is reabsorbed here by active transport. Calcium reabsorption is activated by parathyroid hormone (PTH).
Distal part:
The permeability of the distal part of the DCT depends on the presence of two hormones. Aldosterone acts on so-called principal cells. It has two effects:
- It increases the production of a sodium-chloride symporter, which increases reabsorption of sodium and chloride.
- It increases the secretion of K+
Anti-diuretic hormone (ADH) increases the production of a type of water channel called aquaporin 2 (AQP2), which increases reabsorption of water.
Collecting tubule
Aldosterone and ADH increase sodium, chloride and water reabsorption in the collecting tubule, too.
The collecting tubule has a type of cell called intercalated cells. These cells are essential for pH regulation as they can secrete or reabsorb H+ or HCO3– depending on what the body needs to maintain normal pH. Aldosterone acts on these intercalated cells and stimulates H+ secretion.