Table of Contents
Page created on November 28, 2018. Last updated on December 13, 2018 at 14:48
Microbial carcinogenesis
Three types of microbes can be cancerous, RNA viruses, DNA viruses and bacteria. The most important are:
- RNA virus
- HTLV-1 (Human T-lymphotropic virus 1)
- HCV (Hepatitis C virus)
- DNA virus
- EBV (Epstein-Barr virus)
- HPV (Human papillomavirus)
- HBV (Hepatitis B virus)
- HHV8 (Kaposi sarcoma virus)
- Merkel cell polyoma virus
- Bacteria
- Helicobacter pylori
RNA viruses
HTLV-1 causes T-cell leukaemias and lymphomas and is endemic in Japan and the Caribbean Islands. Like HIV it targets CD4+ cells (HIV is also called HTLV-3), and also like HIV the primary infection lies latent for a long time (40-60 years!). Infection occurs after sexy sex, blood transfusion or breast feeding. The process of carcinogenesis of the virus is unknown. The genome of HTLV-1 contains a gene called TAX which is important for the carcinogenesis.
Hepatitis C and hepatitis B together account for more than 70% of all hepatocellular carcinoma cases. HCV and HBV have a similar, multifactorial carcinogenic process:
- Infection causes an immune-mediated chronic inflammation
- Hepatocytes die
- Hepatocytes regenerate, and NF-κB is activated
- NF-κB blocks apoptosis, allowing hepatocytes to accumulate genetic damage
- Cirrhosis occurs
- Hepatocellular carcinoma occurs
In addition to this immune-mediated carcinogenesis can the HCV virus contain genes that are cancer-promoting.
DNA viruses
Many types of human papilloma virus exist, at least 70 genotypes. Some of them are low-risk, having only a low chance of promoting cancer, while others are high-risk.
Low-risk genotypes cause:
- Benign squamous lesions, like papillomas or warts
- Lesions with low malignant potential, like CIN I
High-risk genotypes cause malignant transformation when the virus integrates its genome into the cell of the host. The site of integration is random, meaning that it can integrate anywhere in the genome. The cells that are infected like this show genomic instability, meaning that they accumulate mutations much easier than healthy cells. The result is usually squamous cell carcinoma in the cervix, anogenital region or oropharyngeal region.
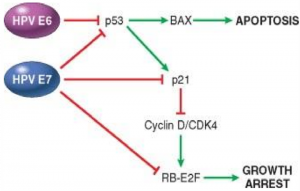
HPV virus has two important genes, E6 and E7, and both code for proteins with the same name. E6 binds to and inhibits p53, while E7 binds to and inhibits pRb, p21, p53 and p27. Cell culture studies have shown that infection with HPV is not by itself enough to form cancer. Cell cultures infected with HPV and with RAS mutations become malignant.
Epstein-Barr virus is a DNA virus that is associated with many types of cancer, like:
- Burkitt lymphoma
- B-cell lymphomas (in immunodeficient hosts)
- Hodgkin’s lymphoma
- Nasopharyngeal carcinoma
- Gastric carcinomas
- Rare NK/T cell lymphomas
EBV infects B-cells by using the complement receptor 2 (CD21). The virus doesn’t replicate itself in the cell, nor does it cause cell necrosis. Instead it inhibits the B-cells’ ability to undergo apoptosis, while simultaneously tricking it into believing it’s being stimulated constantly. EBV encodes for two oncogenes:
- LMP1 activates NF-κB and JAK/STAT signalling, which tricks the cell into believing that its CD40 co-receptor is being stimulated
- EBNA2 activates cyclin D, tricking the cell into believing its Notch receptor is being stimulated
The result is that the EBV-infected cells become immortal and start proliferating, which causes a large polyclonal EBV-infected B-cell population. This is seen clinically as infectious mononucleosis, often just called mono or kissing disease.
Cancer still hasn’t developed. It only develops once the c-myc oncogene becomes involved, often by translocations. Common translocations include:
- t(8;14)
- t(8;22)
- t(2;8)
All these translocations cause Burkitt lymphoma, an aggressive B-cell lymphoma that is very common in childhood in central Africa. 90-100% of all Burkitt lymphoma cases in Africa are associated with EBV, but in Europe only 15-20% of cases are.
Nasopharyngeal carcinoma is endemic in south China and some parts of Africa. EBV-infected cells are found in all tumors of this type. It’s unknown how the virus can enter the epithelial cells as they don’t have the CD21 surface protein. The pathomechanism is similar as for B-cells, where the virus stimulates NF-κB pathway and pro-angiogenic factors.
Hepatitis B has a similar cancer-promoting mechanism as hepatitis C. HBV is endemic in far east and Africa countries, where the incidence of hepatocellular carcinoma is the highest in the world.
Helicobacter pylori
H. pylori is most known for its ability to form gastric ulcers, but these ulcers can be taken a step further to produce gastric adenocarcinomas and MALT lymphomas.
The pathomechanism of adenocarcinomas is similar to that of the hepatitis viruses. It begins with a chronic inflammation (gastritis) which leads to gastric atrophy, intestinal metaplasia of the epithelium, dysplasia and finally cancer. The process takes decades and affects only 3% of infected people.
In addition to the inflammation-induced process does the bacterium itself produce a toxin called CagA. The toxin is injected into the gastric epithelial cells, which initiates a signal cascade that mimics unregulated growth factor stimulation.
The pathomechanism of MALT lymphomas isn’t as well understood. It probably begins with B-cells in the MALT being activated by bacterial antigens, causing them to proliferate. One of the proliferating B-cells accumulate mutations, creating a monoclinal MALToma.
Hey,
The E7 oncogene in HPV inhibits pRB, p21 and p27. That what they say in the lectures
Fixed, thanks.
Hi,
the professor mentioned that HTLV-1 is very similar to its “brother” HTLV-3(HIV). This is a nice way to remember ways of infection and that it attacks CD4+ cells.
That’s a nice rule to remember. I’ll add that in, thanks!
Does the sex that causes HTLV-1 have to be sexy or can it be relatively average? Asking for a friend
It has to be sexy 😏 You can tell your friend to call me ☎
😂😂😂